
مدیتیشن ذهن آگاهی با تغییرات طولانیمدت در توپولوژی عملکردی هیپوکامپ در حالت استراحت ارتباط دارد: پژوهش مغناطیس آنفالوگرافی
خلاصه پژوهش
پیشنهاد شده است که تمرین مدیتیشن با پدیدههای نوروپلاستیسیته (انعطافپذیری عصبی)، کاهش انحطاط مغزی مرتبط با سن و بهبود عملکردهای شناختی مرتبط است. مطالعات تصویرسازی عصبی نشان میدهد که ارتباطات مغزی در مدیتیتورها تغییر میکند. در پژوهش حاضر، هدف ما توصیف اثرات طولانیمدت احتمالی مدیتیشن بر شبکههای مغزی بوده است. برای این منظور، ما از مغناطیس آنفالوگرافی برای مطالعه شبکههای عملکردی مغز در حالت استراحت در مراقبه کنندگان ویپاسانا استفاده کردیم و تغییرات توپولوژیکی شبکه مغزی را در مدیتیتورها در مقایسه با گروه شاهد مشاهده کردیم. بهطور خاص، در باند تتا، مدیتیتورها ازنظر آماری درجه بالاتری (009/0 p = تصحیح شده) درجه بالاتری (شاخص مرکزیتی که نشاندهنده تعداد اتصالات ناشی از یک گره معین است) در هیپوکامپ راست را در مقایسه با گروه شاهد نشان دادند. با در نظر گرفتن نقش هیپوکامپ در فرایندهای حافظه و در پاتوفیزیولوژی بیماری آلزایمر، مدیتیشن ممکن است نقش بالقوهای در حوزه استراتژیهای پیشگیرانه داشته باشد.
معرفی پژوهش مغناطیس آنفالوگرافی
خلاصه
پیشنهاد شده است که تمرین مدیتیشن با پدیدههای نوروپلاستیسیته (انعطافپذیری عصبی)، کاهش انحطاط مغزی مرتبط با سن و بهبود عملکردهای شناختی مرتبط است. مطالعات تصویرسازی عصبی نشان میدهد که ارتباطات مغزی در مدیتیتورها تغییر میکند. در پژوهش حاضر، هدف ما توصیف اثرات طولانیمدت احتمالی مدیتیشن بر شبکههای مغزی بوده است. برای این منظور، ما از مغناطیس آنفالوگرافی برای مطالعه شبکههای عملکردی مغز در حالت استراحت در مراقبه کنندگان ویپاسانا استفاده کردیم و تغییرات توپولوژیکی شبکه مغزی را در مدیتیتورها در مقایسه با گروه شاهد مشاهده کردیم. بهطور خاص، در باند تتا، مدیتیتورها ازنظر آماری درجه بالاتری (009/0 p = تصحیح شده) درجه بالاتری (شاخص مرکزیتی که نشاندهنده تعداد اتصالات ناشی از یک گره معین است) در هیپوکامپ راست را در مقایسه با گروه شاهد نشان دادند. با در نظر گرفتن نقش هیپوکامپ در فرایندهای حافظه و در پاتوفیزیولوژی بیماری آلزایمر، مدیتیشن ممکن است نقش بالقوهای در حوزه استراتژیهای پیشگیرانه داشته باشد.
توضیحات پژوهش پیرامون مدیتیشن ذهن آگاهی
نتایج چندین مطالعه نشان داده است که تمرین مداوم مراقبه یا مدیتیشن باعث ایجاد پدیدههای پلاستیسیته عصبی از جمله کاهش انحطاط مغزی مرتبط با سن و بهبود عملکردهای شناختی میشود. همچنین، بهطور خاص، تأثیرات مدیتیشن با بهبود توجه، حافظه فعال، تواناییهای فضایی و حافظه بلندمدت نیز ارتباط دارد.
روشهای مدیتیشن مختلفی وجود دارد که شباهتهایی با یکدیگر دارند اما در شیوهها و جهتگیری نظری متفاوت هستند. اخیراً، تمرینات مدیتیشن به دو دسته کلی مراقبه متمرکز (سبک متمرکز concentrative style) و مدیتیشن آگاهی باز (تمرکز حواس mindfulness) تقسیم شدهاند. بسته به نحوه هدایت فرآیندهای توجه، تکنیکهای مدیتیشن متمرکز شامل تمرکز بر فعالیتهای ذهنی یا حسی خاص، مانند تکرار صدا، تصویر ذهنی یا احساسات بدنی خاص مانند تنفس است. از سوی دیگر ، ویژگی اساسی مدیتیشن ذهن آگاهی نوع خاصی از توجه است که با تفکر بدون قضاوت مشخص میشود و به مدیتیتورها اجازه میدهد در زندگی روزمره به شیوهای “غیر واکنشی” عمل کنند.
طی ده سال گذشته، مطالعات علمی روی یکی از اصلیترین مدیتیشنهای آگاهی باز یا تمرکز حواس ، به نام مدیتیشن ویپاسانا (VM) تمرکز کردهاند. VM یک عمل بودایی است که شامل تمرکز آگاهی حسی از لحظه با شرایط روانی آرام و بدون واکنش است. در زبان هند باستانی پالی، ویپاسانا به معنای “دروننگری، بینش نافذ، مشاهده و درک واقعیت آنگونه که واقعاً هست”، است. با شناخت فرآیندهای ذهنی و احساسی که در طول مدیتیشن رخ میدهد فرد میتواند از طریق VM، به فرا آگاهی برسد.
تصویربرداری عصبی و مطالعات الکتروانسفالوگرافی نشان میدهد که ارتباطات مغزی افراد مدیتیتور در زمان مدیتیشن و همچنین، در حالت استراحت تغییر میکند. علاوه بر این، تمرین مداوم مدیتیشن باعث تغییرات طولانیمدت گستردهای در اتصال ساختاری مغز میشود که نشاندهنده این است که احتمالاً مدیتیشن باعث انعطافپذیری عصبی نیز میشود.
برای مطالعه فعالیت مغز از مگنتوانسفالوگرافی (MEG) استفاده میشود؛ زیرا میتواند مستقیماً نوسانات عصبی مغز را ضبط کند. با حفظ وضوح زمانی بالای EEG، سیگنالهای MEG توسط لایههای اطراف مغز مخدوش نمیشوند و این امر باعث بازسازی دقیق زمانی و مکانی فعالیت عصبی در مغز میشود. دور-زیدرمن و همکاران با استفاده از MEG دریافتند که بین نوع متمایز خودآگاهی که از طریق مدیتیشن ذهن آگاهی به دست میآید و کاهش قدرت نوارهای گاما و بتا در چندین ناحیه خاص از مغز ارتباط معناداری وجود دارد. علاوه بر این، سیگنالهای MEG (و همچنین EEG) امکان استفاده از مراحل را بهمنظور کمیسازی میزان همگامسازی بین مناطق مغز را فراهم میکند.
در مطالعات سالهای اخیر، رویکرد نظریه گراف بهمنظور استخراج ویژگیهای مربوط به فعلوانفعالات بین نواحی مغز برای سیگنالهای عملکردی نیز مورداستفاده قرار گرفته است. مارزتی و همکاران با استفاده از نظریه شبکه برای سیگنالهای MEG دریافتند که مدیتیشن باعث اتصال جفت قشر سینگولی خلفی به گرههای شبکه حالت پیشفرض و شبکه مغزی کنترل اجرایی در محدوده فرکانس آلفا میشود. همچنین، نتایج اخیر یک مطالعه EEG، نشان داده است که مدیتیشن با افزایش ادغام شبکه مغزی نیز همراه است.
علیرغم نقش مستند عمل مدیتیشن بر عملکرد مغز، تابهحال توجه کمی به تأثیرات طولانیمدت تمرین مدیتیشن بر همگامسازی بین مجموعههای عصبی و توپولوژی مغز شده است. تا آنجا که ما میدانیم، هیچ مطالعه MEG پیشرفتهای که همزمانسازی شبکه مغزی و ویژگیهای توپولوژیکی آن را در رابطه با تمرین مراقبه موردبررسی قرار دهد، تاکنون انجامنشده است.
در پژوهش حاضر، فرض بر این بوده است که تمرین معمول مدیتیشن ذهن آگاهی ممکن است اثرات طولانیمدتی در توپولوژی مناطق مغزی درگیر در توجه و/یا حافظه ایجاد کند. برای آزمایش فرضیه مذکور، ما منابع درون مغز را بهعنوان گره و فعلوانفعالات عملکردی بین آنها را بهعنوان رابط در نظر گرفتهایم. همچنین، رابط را بر اساس شاخص تأخیر فاز (PLI) که هماهنگسازی فاز بین مناطق مغزی را مشخص میکند، تعیین کردیم.
متعاقباً، ما حداقل Spanning-tree (MST) را به دست آوردیم که به ما اجازه داد یک زیرگراف منحصربهفرد ایجاد کنیم تا تمام گرههای تشریحی را به هم متصل کند و ستون فقرات اصلی شبکه را مشخص کند. همچنین، وضعیت شبکه مغزی در گروهی از مدیتیتور های معمول Vipassana و گروههای کنترل (افرادی که تجربه مدیتیشن نداشتند) در حالت استراحت را از طریق مقایسه توپولوژی تجزیهوتحلیل کردیم.
روش پژوهش:
شرکتکنندگان
این پژوهش بر روی 29 نفر که بیش از یک سال VM کار میکردند و 31 نفر که هرگز در زندگی خود مدیتیشن نکرده بودند، انجامشده است. شرکتکنندگان مدیتیتور از جامعه ویپاسانا کایوانو (ناپل) انتخاب شده بودند. همچنین، همه شرکتکنندگان در این مطالعه در سال گذشته حداقل پنج روز در هفته به مدت 1 ساعت یا بیشتر، مدیتیشن کرده بودهاند.
معیارهای دیگر برای انتخاب شرکتکنندگان به شرح زیر بوده است: عدم ابتلا به هر گونه بیماری داخلی، عدم ابتلا به هرگونه بیماری عصبی یا روانی سابقه شرکت در ویپاسانا در سال گذشته و چپدست نبودن. در میان شرکتکنندگان سه مدیتیتور به دلایل زیر رد شدند: یکی چپدست بود، یکی بیماری عصبی داشت و دیگری به دلیل کیفیت پایین سیگنالهای MEG.. دو نفر نیز به دلیل نویز سیگنال زیاد از گروه کنترل کنار گذاشته شدند.
گروه مدیتیتورهای ویپاسانا متشکل از بیستوشش شرکتکننده (هشت مرد و هجده زن)، با میانگین سنی 42.6 سال (± SE 2.4)، میانگین سالهای تجربه مدیتیشن 6.41 (± SE 1.4) در این پژوهش موردبررسی قرار گرفتند. گروه کنترل نهایی متشکل از بیستونه نفر (نه مرد و بیست زن) در گروههای سنی-جنسیتی-تحصیلی-نژادی، میانگین سنی 43 سال (1. SE 1.98) ،که هیچ تجربهای در مدیتیشن نداشتند بوده است. همه شرکتکنندگان حاضر در این پژوهش بومیان ایتالیایی بودند (جدول 1)
|
جدول 1 ویژگی کوهرت، مفهوم و خطای استاندارد گزارش شده است. |
||||||
|
ویژگیهای گروه |
سایز |
جنسیت(مرد/زن) |
سن |
تحصیلات |
سابقه مدیتیشن |
ساعتهای مدیتیشن/روز |
|
مدیتیتور کنترل |
26 29 |
18/8 20/9 |
42.30 ± 2.4 43 ± 1.98 |
15.12 ± 0.66 15.72 ± 0.70 |
6.4 ± 1.4 – |
1.33 ± 0.72 – |
این مطالعه مطابق توصیههای Comitato Etico Campania Centro انجام شده است. این پروتکل توسط “Comitato Etico Campania Centro” (Prot.n.93C.E./Reg. n.14-17OSS) تأیید شده است. تمام شرکتکنندگان مطابق با اعلامیه هلسینکی رضایتنامه کتبی خود را از شرکت در این پژوهش اعلام کردند.
حصول MEG
سیستم MEG توسط شورای تحقیقات ملی، Pozzuoli، ناپل، در موسسه علوم کاربردی و سیستمهای هوشمند “E. کایانیلو. توسعه یافته است.
این سیستم مجهز به 163 مغناطیسسنج شامل 9 سنسور مرجع است که در فاصله بیشتری از کلاه ایمنی قرار دارد و در یک اتاق محافظ مغناطیسی (ATB ،Ulm، آلمان) قرار انجام میشود تا نویز پسزمینه را کاهش دهد. مالکیت، پیشپردازش و بازسازی منبع همانطور که قبلاً توضیح داده شد انجام شده است. جزئیات بیشتر، قبل از حصول، چهار حلقه موقعیت روی سر سوژه قرار داده شده است و با استفاده از Fastrak (Polhemus®) دیجیتالی شده است. کویل ها در ابتدای هر بخش از ثبت فعال میشوند. سوژهها روی یک صندلی راحتی در اتاق محافظ نشستند. سیگنالهای الکتروکاردیوگرافی و الکتروکولوگرافی برای کمک به حذف مصنوعات باهم ضبط میشوند. فعالیت مغز (شکل 1 (الف)) در حالت استراحت در دو بخش مجزا به مدت 2.5 دقیقه با چشمان بسته ثبت شدند. دادههای MEG، پس از فیلتر ضد نفوذ، با فرکانس نمونهبرداری 1024 هرتز به دست آمد. سپس، سیگنالها با استفاده از فیلتر درجه چهارم Butterworth IIR در باند 0.5-48 هرتز فیلتر شد.
تصویر شماتیک بازسازی سیگنالهای MEG. الف) سیگنال های MEG. (ب) پرتو سازی بر اساس MRI الگو. (ج) سریهای زمانی بازسازیشده برای 90 منطقه موردعلاقه (ROI). (د) ماتریس اتصال شامل اتصالات عملکردی بین ROI بر اساس PLI. ه) ساخت شبکه مغزی مبتنی بر MST (که در آن هر ROI یک گره و هر اتصال عملکردی یک لبه است).
پیشپردازش
در مرحله اول، از تجزیهوتحلیل اجزای اصلی (PCA) برای کاهش سروصدای محیط استفاده شد. بهطور خاص، فیلتر با متعامد سازی سیگنالهای مرجع برای به دست آوردن یکپایه، نمایش سنسورهای مغزی بر اساس نویز و حذف پیشبینیها برای به دست آوردن دادههای بینقص به دست آمد. ما برای این کار از پیادهسازی فیلتر PCA موجود در جعبهابزار Fieldtrip استفاده کردیم. متعاقباً، کانالهای پر سروصدا بهصورت دستی از طریق بازرسی بصری توسط یک رتبهبندی باتجربه حذف شدند. سرانجام، برای هر موضوع، تجزیهوتحلیل اجزای مستقل (ICA) برای حذف مصنوعات فیزیولوژیکی، قلبی (بهطورکلی یک جزء) و چشمک زدن (در صورت وجود) از سیگنالهای MEG استفاده شد.
برای هر موضوعی که شامل مصنوعات (اعم از سیستم یا فیزیولوژیکی) یا صدای محیطی قوی نبودند ده دوره 8 ثانیهای انتخاب شد. طول 8 ثانیه برای جبران نیاز به داشتن دورههای مشخص بهاندازه کافی، برای جلوگیری از خوابآلودگی و به دست آوردن برآورد قابلاطمینان از میزان ارتباطات است.
بازسازی منبع
تمام پردازشهای مربوط به روش ایجاد پرتو با استفاده از جعبهابزار Fieldtrip انجام شده است. بر اساس الگوی MRI ، مدل هدایت حجمی پیشنهاد شده توسط نولته در نظر گرفته شده است و حداقل واریانس محدودیت خطی (LCMV) برای بازسازی سریهای زمانی مربوط به سانتریوئیدهای 116 منطقه موردعلاقه (ROIs) اجرا شده است. ). اگرچه نواحی مخچهای قبلاً در برخی از مطالعات EEG بازسازی شده بودند، اما ما تصمیم گرفتیم آنها را (با در نظر گرفتن 90 ROI) با توجه به قابلیت اطمینان پایینتر ، وارد این پژوهش نکنیم. برای هر منبع، ما سری زمانی را در امتداد دوقطبی که بیشترین واریانس را با تجزیه مقدار واحد (SVD) مشخص میکردند، پیشبینی کردیم. برای سریهای زمانی منبع در 512 هرتز مجدداً نمونه انجام شدند (شکل 1 (ج)).
2.5 اتصال و تجزیهوتحلیل شبکه
برای برآورد اتصال عملکردی PLI، از نرمافزار BrainWave استفاده شده است (CJS، نسخه 09.152.1.23 بر اساس توزیع تفاوت فازهای لحظهای (∆Φ (t)) (ناشی از تبدیل هیلبرت سری زمانی) و برای سریهای دو زمانه است و بهصورت زیر محاسبه میشود:
PLI=|⟨sign[sin(ΔΦ(tk))]⟩|,
جایی که “<>” مقدار متوسط را نشان میدهد، “علامت” مخفف تابع signum است، “|. |” مقدار مطلق را نشان میدهد و “tk” نمونهها هستند. تفاوت فاز در محدوده [−π ، π] تعریف شده است. این اندازهگیری نسبت به هدایت حجم حساس نیست. مقادیر PLI بین 0 و 1 است که در آن 1 نشاندهنده همگامسازی کامل و 0 نشاندهنده فعالیت غیر همزمان است.
ما یک ماتریس مجاورت 90 × 90 را برای هر دوره هر موضوع، در همه باندهای فرکانسی به دست آوردیم. برای هر دوره، ماتریس PLI محاسبه شد و پسازاین مرحله، آنها با میانگین حسابی ادغام شدند. بنابراین، با محاسبه PLI برای هر دو ناحیه مغزی، ماتریس مجاورت وزنی 90 90 90 (شکل 1 (د)) را برای هر دوره و برای هر موضوع، در همه باندهای فرکانسی به دست آوردیم: دلتا (0.5-4.0 هرتز )، تتا (4.0-8.0 هرتز)، آلفا (8.0-13.0 هرتز)، بتا (13.0-30-30 هرتز) و گاما (30.0-48.0 هرتز).
ماتریس مجاورت وزنی برای بازسازی شبکه مورداستفاده قرار گرفت، جایی که 90 ناحیه اطلس AAL بهعنوان گره نشان داده شده و مقادیر PLI لبههای وزنی را تشکیل داد. همچنین، حداقل spanning tree مخصوص فرکانس نیز برای هر دوره محاسبه شد. (شکل 1 (ه)). ازآنجاکه ما علاقهمند به ارتباطات قوی بودیم، برای ساخت MST، وزن لبه 1/PLI نیز تعریف شد. در واقع، الگوریتم کروسکال [48] ابتدا لینکها را به ترتیب صعودی رتبهبندی کرد و سپس، شبکه را با افزودن یک لینک در یکزمان و دور انداختن لینکهایی که لوپ ایجاد میکنند، تعریف کرد. الگوریتمها تا زمانی ادامه پیدا میکرد که همه گرهها به هم متصل شوند و در نتیجه، یک نمودار بدون حلقه با گرههای N و لینکهای M = N – 1 ایجاد شود.
برای جلوگیری از برخی از سوگیریها در تجزیهوتحلیل شبکههای سنتی، ما از حداقل spanning tree (MST) استفاده کردیم که تفسیر توپولوژیکی بیطرفانهای از نتایج را ممکن سازد. بر اساس MST، هم پارامترهای جهانی (کسر برگ، واگرایی درجه و سلسله مراتب درختان) و هم پارامترهای گرهای (درجه، مرکزیت بین مرکزیت و گریز از مرکز) را محاسبه کردیم. کسر برگ بهعنوان کسری از گرهها با درجه 1 تعریف میشود که نشاندهنده یکپارچگی شبکه است.
کسر برگ بالاتر نشاندهنده این است که شبکه به سمت توپولوژی ستارهای تمایل دارد و جایی که گرهها بهطور متوسط در مقایسه با توپولوژی خطیتر به یکدیگر نزدیکتر هستند واگرایی درجه، معیاری از گستردگی توزیع درجه، مربوط به تابآوری در برابر حملات است. سرانجام، سلسلهمراتب درختی بهعنوان تعداد برگ بر حداکثر مرکزیت بین حدفاصل تعریف شده است. ایده پشت این اندازهگیری این بوده است که یک شبکه بهینه باید از ارتباطی مؤثر و در عین اجتناب از اضافهبار هاب برخوردار شود. سلسلهمراتب درختی نیز بهمنظور تعیین تعادل بین هر دو ویژگی طراحی شده است.
علاوه بر این، بهمنظور تعیین اینکه آیا مناطق خاصی با توجه به سالهای مدیتیشن تفاوت دارند یا خیر ما درجه مرکزیت بین مرکز (BC) و گریز از مرکز را برای هر گره محاسبه کردیم.
BC بهعنوان تعداد کوتاهترین مسیرهایی که از یک گره معین درمجموع کوتاهترین مسیرهای شبکه عبور میکنند تعریف میشود. گریز از مرکز نیز بهعنوان طولانیترین مسیر بین یک گره و گره دیگر شبکه تعریف میشود. هرچه گریز از مرکز کمتر باشد، گره مرکزیتر است. این معیارها برای هر دوره محاسبه شده است و متعاقباً در هر دوره بهطور میانگین برای هر موضوع بهطور متوسط محاسبه شده است.
تحلیل آماری
تمام تجزیهوتحلیلهای آماری در Matlab (Mathworks®، نسخه R2013a) انجام شده است. برای مقایسه دو گروه، در هر باند فرکانسی و برای هر پارامتر، از آزمون جایگشت استفاده کردیم که در آن توزیع صفر بهعنوان تفاوت بین گروه دادهها بهدستآمده است. هر بار، تفاوت میانگینهای دو گروه محاسبه شده است و توزیع صفر برای تفاوت بین گروهها به دست آمد که از چنین توزیعی برای تعریف اهمیت آماری تفاوت مشاهده شده بین دو گروه استفاده شد. ما از نرخ کشف کاذب (FDR) برای تصحیح مقایسههای متعدد استفاده کردیم و در نهایت، سطح معنیداری 05 /0 ≤ p تعیین گردیده است.
نتایج
پس از ایجاد یک ماتریس مجاورت مبتنی بر PLI، ما از یک روش شبکه مبتنی بر MST برای مقایسه پارامترهای توپولوژیکی گرهای و جهانی در مراقبهها و کنترلها استفاده کردیم.
پارامترهای جهانی (کسر برگ، واگرایی درجه و سلسلهمراتب درخت) قبل از تصحیح برای مقایسههای متعدد بین معیارها، بین گروهها تفاوت معنیداری از خود نشان نداد.
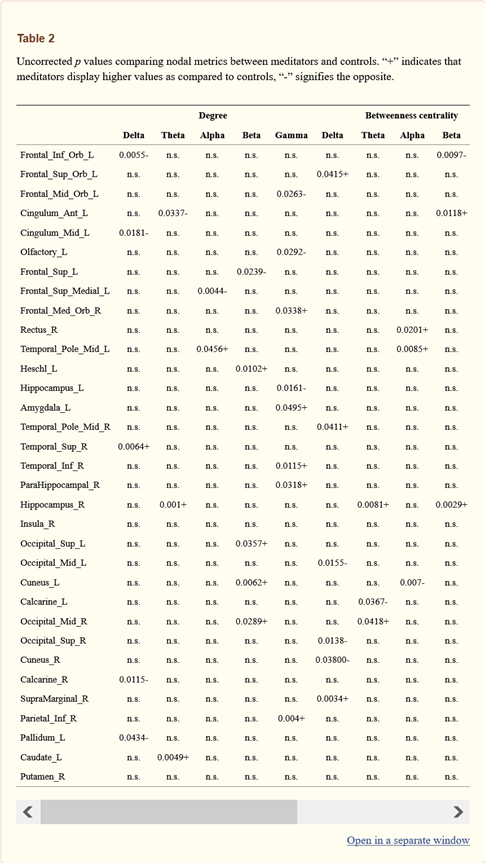
با توجه به معیارهای گره، قبل از اصلاح در مقایسههای متعدد ، تفاوتهایی بین مراقبه کنندگان و گروه کنترل در مناطقی که معمولاً تحت تأثیر تمرین مراقبه قرار دارند، مشاهده شد. بهطور خاص، در آمیگدال، در باند گاما، درجه و BC در مراقبه کنندگان در مقایسه با گروه کنترل بیشتر بود (به ترتیب 0.049 = p ، 0.034 = p). caudatus، ناحیه دیگری که تحت تأثیر VM قرار دارد، نشان داده شده است که در مدیتیتورها در نوار تتا بیشتر متصل است (0049 = p). در قشر پیش پیشانی، تفاوتهای نسبتاً گستردهای را در مناطقی مانند جایروس رکتس (در میان سایر موارد) gyrus rectus در نوار آلفا (02012 = p) و سینگولوم در نوار تتا (0181 = p) پیدا کردیم. قشر کالکارین در باند تتا (0.011 = p) کمتر و در باند گاما (0.038 = p) متصلتر به نظر میرسید. برای تفسیر نتایج، مقادیر p اصلاحنشده را در جدول 2 گزارش کردیم. با اینحال، نکته مهم این است که ازآنجاییکه این نتایج FDR تصحیحنشده است باید بااحتیاط شدید تفسیر شود.
مقادیر p اصلاحنشده در مقایسه معیارهای گره بین مراقبه کنندگان و کنترلها.
“+” نشان میدهد که مدیتیتورها مقادیر بالاتری را در مقایسه با کنترلها نشان میدهند و “-” نتیجه برعکس را نشان میدهد.
پس از تصحیح مقایسههای متعدد بین مناطق، در مدیتیتورها درجه بالاتری (009/0 p = تصحیح شده) در نوار تتا، در هیپوکامپ راست (شکل 2) مشاهده شد. هیچ اهمیت گرهای دیگری FDR را در سایر باندهای فرکانسی مشاهده نشد.
شکل 2
میانگین درجه در نوار تتا در هیپوکامپ راست.
دادهها بهصورت میانگین. SD بیان شده است. ستاره نشاندهنده معنیداری آماری p = 0.009 است.
مطالعه و بررسی تخصصی
این مطالعه بهمنظور بررسی طولانیمدت مدیتیشن ذهن آگاهی در ویژگیهای توپولوژیکی در شبکههای مغزی انجام شده است. برای این منظور، ما از تجزیهوتحلیل شبکه برای مقایسه دادههای MEG بهدستآمده از گروهی از مدیتیتورها Vipassana با کنترلها، در شرایط حالت استراحت استفاده کردهایم. هیچ تفاوتی در معیارهای جهانی مشاهده نشد. علاوه بر این، قبل از اصلاح FDR، ما توپولوژی تغییریافتهای را در مدیتیتورها در تعدادی از نواحی مغز ازجمله لوبهای جلویی، لوبهای گیجگاهی، لوبهای اکسیپیتال و آمیگدال مشاهده کردیم، تصحیح FDR، نتایج ما نشان داد که هیپوکامپ راست در مقایسه با گروه کنترل درجه بالاتری را در باند تتا نشان میدهد.
همانطور که انتظار میرفت، ما در مقایسه بین مدیتیتورها و گروه کنترل در معیارهای جهانی تفاوتی پیدا نکردیم. معیارهای جهانی سازماندهی مجدد گستردهای را در فعالیت مغزی نشان میدهد و مشخص شد که این خاصیت معمولاً در فرآیندهایی که باعث آسیبهای منتشر به مغز میشوند، مانند بیماریهای عصبی، تغییر میکند. نتایج ما نشان داد که اتصال یا شبکه مغزی (کسر برگ)، مقیاس آزاد بودن آن (واگرایی درجه) و انعطافپذیری آن در برابر حملات هدفمند (سلسلهمراتب درختان) تحت تأثیر VM قرار نمیگیرند. از اینرو، میتوان نتیجه گرفت که مدیتیشن باعث سازماندهی مجدد در کل شبکه نمیشود.
با توجه به نتایج گرهای که برای مقایسههای متعدد تصحیح نشده است، طبق ادبیات قبلی، ما مناطقی را با توپولوژی تغییریافته در لوبهای جلویی مشاهده کردیم، در لوبهای گیجگاهی، در آمیگدال، در caudatus، و در لوبهای پسسری در چند باند فرکانسی. نتایجی که ما پیدا کردیم در چند باند فرکانسی بود که بهطورکلی ثبتشده بود. با اینحال، به دلیل عدماصلاح مقایسههای متعدد، چنین نتایجی صرفاً اکتشافی محسوب میشود.
پس از اصلاح چندین مقایسه بین مناطق، هیپوکامپ راست مدیتیتورها درجه بالاتری را در نوار تتا نشان دادند. این نتیجه مطابق با شواهد قبلیای است که نقش مرکزی هیپوکامپ را در تمرینات مراقبه نشان میدهد. درجه بالاتر نشان میدهد که مدیتیتورها در مقایسه با گروه شاهد بیشتر دچار فعلیت هیپوکامپ راست میشوند.
از این حالت، میتوان نتیجه گرفت که هیپوکامپ راست بیشتر به بقیه شبکه متصل است، به عبارت دقیقتر، این بدان معناست که توپولوژی ناحیه (یعنی خواص روابط منطقی آن با بقیه مغز) بهگونهای اصلاح شده است که نقش آن ناحیه در شبکه مغز بیشتر برجسته شود. به شرطی که در حال بررسی تأثیرات طولانیمدت مدیتیشن باشیم، میتوان انتظار داشت که مناطقی بیشتر در تمرین مراقبه دخیل هستند که تحت تغییرات عملکردی طولانیمدتی قرار دارند. با توجه به این شواهد، مشاهده شد که مدیتیتورها حجم بیشتری و غلظت ماده خاکستری بیشتری را در هیپوکامپ راست در مقایسه با افراد غیر مدیتیتور نشان میدهند. نکته جالب اینجاست که ارتباط همان منطقه در حین تمرین مدیتیشن نیز افزایش بیشتری نشان میدهد. چنین نتایجی ممکن است با یافتههای ما مطابقت داشته باشد و نشاندهنده این باشد که تغییرات بلندمدت توسط VM در هیپوکامپ راست ایجاد میشود.
ازآنجاییکه نتایج ما در این پژوهش مخصوص هیپوکامپ راست است، ممکن است نشاندهنده عملکردی متفاوت بین هیپوکامپ راست و چپ باشد. در حالیکه پیش از این مشاهده شده است که هیپوکامپ چپ هم در ساخت رویدادهای گذشته و هم در رویدادهای آینده فعال است و هیپوکامپ راست فقط در هنگام ایجاد رویدادهای آینده (حافظه آیندهنگر) فعال میشود. همچنین، شواهد علمی نشان میدهد که هنگام پردازش اطلاعات جدید، همگامسازی بین هیپوکامپ و قشر پیش پیشانی عمدتاً در نوار تتا اتفاق میافتد. از اینرو، در طول تخیل وقایع آینده، حافظه جدید و عناصر جدید آموخته میشوند و حافظه آیندهنگر ساخته میشود. علاوه بر دخالت در حافظه آیندهنگر، هیپوکامپ راست در فرآیندهای حافظه دیداری-فضایی نیز دخیل است.
در نهایت، مشاهده کردیم که تفاوت مرکزیت هیپوکامپ مختص باند تتا است و همانطور که در بالا ذکر شد، در فرآیندهای حافظه نیز دخیل است. به گفته Buzsáki، نوسانات تتا بهعنوان “سازمان دهنده زمانی ضروری” در نظر گرفته میشود و به جهتگیری زمان در طول حافظه اپیزودیک و همچنین، دنیای فیزیکی کمککننده است. همچنین، در نواحی زمانی، افزایش قدرت تتا در حین تمرین مراقبه بهطور گستردهای گزارش شده است.
با توجه به نقش عملکردی هیپوکامپ راست و فعال شدن آن در نوار تتا، نتایج ما نشان میدهد که مدیتیشن ممکن است تأثیرات عملکردی بر خاطرات آینده و فضایی داشته باشد. این یافتهها نشان میدهد که اگر مدیتیشن، بهطور مداوم انجام شود، میتواند بهعنوان یک مداخله غیر دارویی در آسیبشناسیهایی که با تغییر در مناطق هیپوکامپ مشخص میشوند، محسوب شود.
در نهایت، باید توجه داشت که اغلب دادههای متناقضی نیز وجود دارد که ممکن است به دلیل رویکردهای مختلف روششناختی، مانند تجزیهوتحلیل مرحله مراقبه در مقابل حالت استراحت، نوع مدیتیشن موردبررسی (توجه متمرکز در مقابل مراقبه ذهن آگاهی)، و نحوه دستیابی به دادهها و/یا تجزیهوتحلیل دادهها باشد. نکته قابلتوجه این است که مطالعه ما تنها مطالعهای است که تا به امروز تفاوت شبکه مغزی بین مراقبه کنندگان و کنترلکنندههای ویپاسانا را در حالت استراحت و با استفاده از مگنتوانسفالوگرافی سطح منبع مورد تجزیهوتحلیل قرار داده است.
به نظر میرسد نتایج این پژوهش نویدبخش تائید تأثیرات مدیتیشن بر شبکههای مغزی است. با اینوجود، مهم است که تأکید شود که این مطالعات مقدماتی هستند و نیاز به تجزیهوتحلیلهای بیشتری دارند. بهعنوانمثال، بررسی چشمانداز و حافظه فضایی در مراقبه کنندگان و افراد غیر مدیتیتور در حین انجام وظایف بهمنظور ارتباط اتصال عملکردی با عملکرد شناختی ضروری است. علاوه بر این، تجزیهوتحلیل شبکههای مغزی در افرادی که سایر انواع مدیتیشن را انجام میدهند، نیز میتواند به ما کمک کند تا مزایای شیوههای مختلف مدیتیشن را بهتر درک کنیم.
علیرغم محدودیتهای ذکرشده، نتایج بدست آمده از این پژوهش حاکی از آن است که تمرین مدیتیشن معمول با تغییر طولانیمدت در توپولوژی مناطق مشخص مغز مرتبط است و نشان میدهد که فرآیندها و تاثیرات مراقبه میتواند پلاستیسیته مغز را القا کند.
تقدیر و تشکر
مقاله حاضر توسط دانشگاه Naples Parthenope Ricerca Competitiva 2017 (D.R. 289/2017) به LM و GF و کمکهای مالی از Regione Campania (Sportello Innovazione) به GS انجام شده است. همچنین، از همکاریهای انجمن “پرانا ویدیا یوگا” نیز تشکر میکنیم.
میزان اعتماد و در دسترس بودن دادهها
دادههای مورداستفاده برای حمایت از یافتههای این مطالعه در صورت درخواست از نویسنده مربوطه در دسترس است. بدینوسیله نویسندگان در مورد انتشار این مقاله تائید میکنند که هیچ تضاد منافعی وجود ندارد.
منبع:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6312586/


